系统性红斑狼疮(SLE)是一种由环境、免疫和代谢过程的复杂相互作用引起的严重自身免疫性疾病。树突状细胞(DCs)是有害自身免疫的关键调节者,浆细胞样树突状细胞(pDCs)会产生大量的干扰素,而常规树突状细胞(cDCs)会产生过度的炎症反应。最近观察到,在患有特应性皮炎和银屑病的患者的炎症皮肤以及肿瘤组织中,存在富含免疫调节分子的树突状细胞(mregDCs)的富集,也称为 LAMP3-DCs、CCR7-DCs 或 DC3s。mregDCs 上 CCR7 的高表达表明它们可能是趋化信号的潜在响应者,最近的一项研究表明,mregDCs 会向肿瘤引流淋巴结(dLNs)迁移以抑制 T 细胞免疫。然而,关于控制迁移、存活和功能的代谢机制,以及它们与诸如系统性红斑狼疮等病理自身免疫的关联的了解仍处于初级阶段。
在外周捕获环境信号的迁移性树突状细胞(mig-DCs)在 CCR7 的引导下向引流淋巴结(dLNs)迁移,以引发抗原特异性 T 细胞反应。然而,迁移中的 DCs 如何在自身免疫性疾病的发展过程中协调代谢、信号传导和转录程序来调节细胞存活和随后的适应性免疫,仍然是一个未解决的问题。
本研究中揭示了法尼醇合酶通路(FPP)——胆固醇代谢的中间产物——通过重塑线粒体形态和代谢,在自身免疫性疾病中驱动浆细胞样树突状细胞(mig-DC)功能障碍方面发挥着关键作用。本研究为自身免疫性疾病中 DC 失调的代谢控制提供了见解,并建议了潜在的疾病治疗靶点。
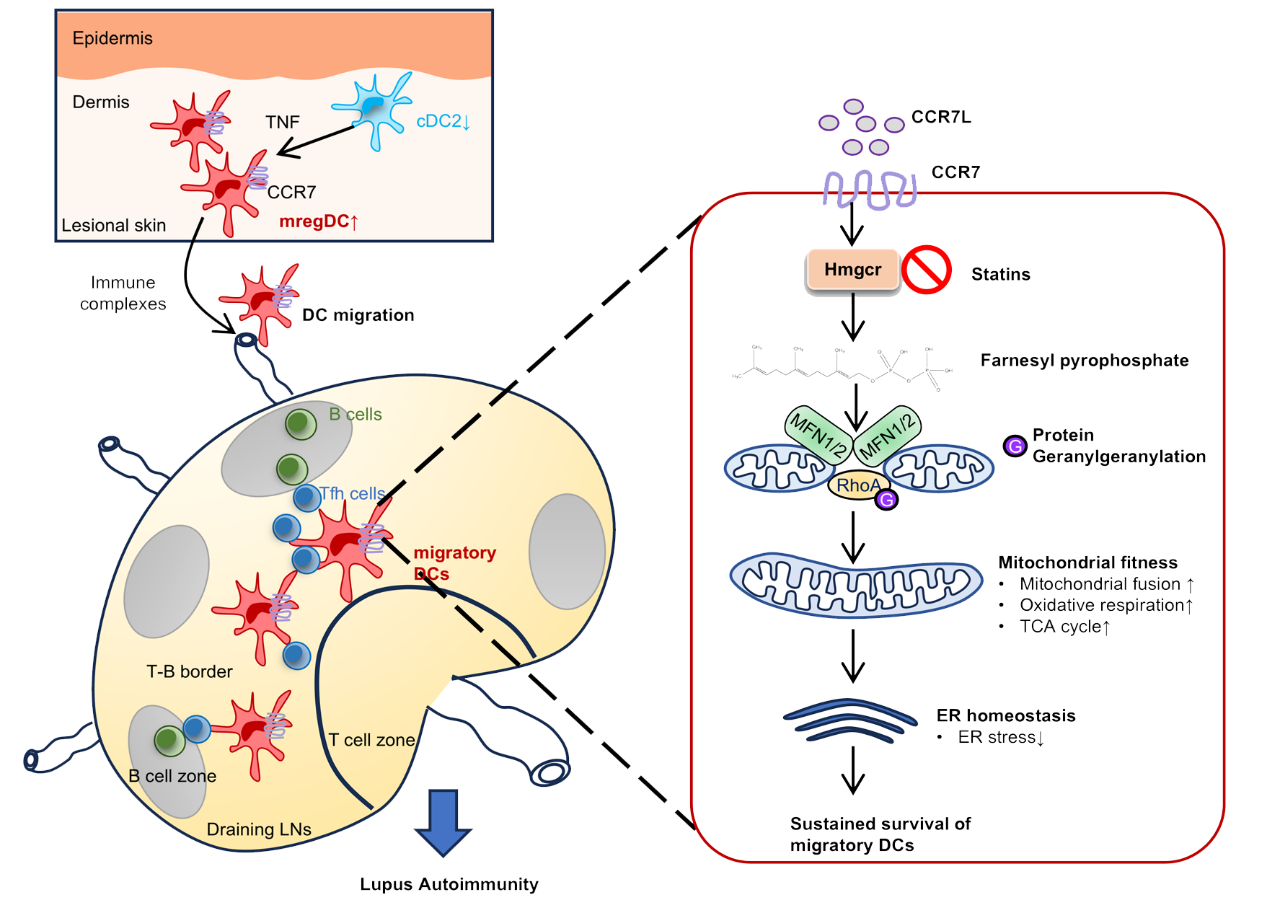
CCR7 介导的树突状细胞迁移在狼疮中增加
皮肤炎症性损伤在狼疮患者中常见,使用公开的单细胞测序数(GSE179633)分析了树突状细胞(DC)亚群的组成和转录特征。该数据集包括 16 个皮肤单细胞悬液。提取了以优先表达 Aif1 和 Lyz 为特征的髓系细胞,并将其进一步分为七个亚簇(SC0 - SC6)。SC5 (CCR7+ DCs)通过表达 mregDC 特征标志物的四个模块与最近鉴定出的 mregDC 相对应,包括(1)DC 激活标志物;(2)调节分子,如 CD274、CD200;(3)迁移或细胞骨架分子,如 CCR7;以及(4)炎性细胞因子,如 CCL17。接下来对 cDC1s、cDC2s 和 mregDCs 进行了轨迹分析。数据表明,cDC2 细胞沿着轨迹逐渐减少,而 mregDCs 逐渐增加,而 cDC1 细胞主要位于轨迹的两端,而非沿着轨迹分布。接着又发现,尽管大多数皮肤中的常规树突状细胞(cDCs)为CD11b+型,但与对照小鼠相比,狼疮小鼠皮肤中CCR7+ cDC2s 的比例更高,而CCR7+ cDC1s 的比例相似。因此,TNF 可能在狼疮自身免疫中促进 cDC2 向CCR7+调节性树突状细胞(mregDCs)的发育。
狼疮巨噬细胞来源的树突状细胞中胆固醇代谢上调
接下来探究在自身免疫性疾病中,是否有任何 CCR7 诱导因子可能导致髓源性调节性树突状细胞(mregDCs)迁移增强。无偏差的 KEGG 富集分析显示,在受到 CCR7L 刺激的髓源性树突状细胞中,类固醇生物合成和萜类骨架生物合成是上调程度最高的前四个通路。一系列编码胆固醇生物合成酶的基因在 CCR7 刺激下显著上调。胆固醇定量测定和菲立磁染色证实了 CCR7 刺激的髓源性树突状细胞中胆固醇的积累。因此,CCR7 刺激在体外诱导了髓源性树突状细胞中胆固醇生物合成通路的激活。
又发现迁移性 DC显示出基因表达模式,包括激活、调节、迁移和炎症分子,与皮肤中的调节性 DC 高度一致,表明淋巴结中的迁移性 DC 是皮肤调节性 DC 的迁移对应物。这表明,亚簇 7(迁移性 DC)高度表达富含 B 细胞激活、T 细胞分化和抗原呈递的基因。在 dLNs 中,通过 Hmgcr 表达和 Bodipy 染色测量的胆固醇水平始终是 mig-DC2s 中最高,高于其他主要的 DC 亚群。
Hmgcr-FPP 通路促进浆细胞样树突状细胞(mDC)的迁移
探究胆固醇通路是否可能对 CCR7 触发的树突状细胞(DC)迁移进行功能性调节。使用脂多糖(LPS)预处理的髓源性 DC 进行体外 Transwell 迁移实验和体内用羧基荧光素二乙酸琥珀酰亚胺酯(CFSE)标记的 DC 迁移实验。使用各种步骤靶向化学物质,我们发现法尼醇(FPP),胆固醇合成途径的中间代谢产物,增强了 CCR7 触发的髓源性 DC 迁移。使用各种步骤靶向化学物质,我们发现法尼醇(FPP),胆固醇合成途径的中间代谢产物,增强了 CCR7 触发的髓源性 DC 迁移。FPP 和胆固醇的联合刺激进一步上调了髓源性 DC 的迁移,表明 FPP 和胆固醇在促进髓源性 DC 迁移方面具有独立的作用。数据证实,在狼疮来源的迁移性树突状细胞中上调的 FPP 在体内选择性地促进 CCR7 触发的髓源性树突状细胞迁移。
FPP 促进免疫复合物诱导的树突状细胞迁移和生发中心反应
由抗原和抗体形成的免疫复合物(ICs)对于狼疮的发病机制至关重要,并且能够刺激自身免疫性疾病中树突状细胞(DCs)的迁移。IC 显著增加了 DC 的迁移,并显著上调了迁移的 DC 中胆固醇的富集。在 OVA-IC 刺激后,辛伐他汀最强烈地减少了髓源性 DC 的迁移,但在 OVA 或 OVA + LPS 刺激后仅适度减少。因此,胆固醇代谢可以在体内平衡、致病和自身免疫的情况下促进髓源性 DC 的迁移,但对于维持由 IC 启动的髓源性 DC 迁移尤为重要。接下来探究了 FPP 在体内促进免疫复合物(IC)启动的髓源性树突状细胞(mDC)迁移中的作用,发现FPP 能够相对挽救由辛伐他汀或 Hmgcr 缺陷引起的缺陷性 DC 迁移。且在 IC 刺激后还减少了深淋巴结中CD4+PD-1+CXCR5+ T 滤泡辅助细胞和GL7+B220+生发中心 B 细胞的比例。因此,FPP 促进了免疫复合物启动的 DC 向深淋巴结 T 细胞 - B 细胞交界区域的迁移,并随后促进了TFH细胞和生发中心反应。
FPP 延长了自身免疫性疾病中迁移性树突状细胞的存活
科学家们试图探究 FPP 在自身免疫性疾病中导致髓源性树突状细胞(mDC)过度迁移的潜在机制。发现在 CCR7 刺激后,Hmgcr 基因敲低或辛伐他汀处理会增加 mDC 的细胞凋亡。因此,Hmgcr - FPP 通路维持了 CCR7 介导的 mDC 存活。FPP 逆转了西伐他汀在 CCR7L 刺激的 mDCs 中引起的促凋亡信号。此外,用 Caspase 3 抑制剂 z-DEVD 抑制细胞凋亡可挽救西伐他汀抑制的 mDC 迁移,而用 Nec-1(坏死抑制剂)、DMF(细胞焦亡抑制剂)或 Fer-1(铁死亡抑制剂)抑制其他形式的程序性细胞死亡则没有此类效果。因此,FPP 通过抑制 Caspase 3 介导的细胞凋亡来促进 CCR7 介导的 mDC 存活和迁移,也发现FPP 延长了 CCR7 介导的迁移性 DCs 的存活。
FPP 促进自身免疫性疾病中巨噬细胞来源的树突状细胞(mig-DCs)的线粒体融合
线粒体是细胞存活和代谢的主要调节者。因此,我们想知道 FPP 是否会影响自身免疫性疾病中巨噬细胞来源的树突状细胞(mig-DCs)的线粒体功能。我们对野生型 mDCs、Hmgcr 缺陷型 mDCs 以及用 FPP 处理的 Hmgcr 缺陷型 mDCs 进行了代谢组学研究,发现由 Hmgcr-FPP 通路调节的代谢物在 TCA 循环中显著富集。多种 TCA 循环中间产物下调,但经 FPP 处理后恢复,如柠檬酸、异柠檬酸、琥珀酸、富马酸和苹果酸。发现 Hmgcr 敲低或缺陷以及辛伐他汀抑制了 CCR7 诱导的线粒体氧化呼吸,而 FPP 处理可挽救这一情况,无论 CCR7 刺激与否,辛伐他汀都会降低浆细胞样树突状细胞(mDCs)的基础呼吸作用,这反映了辛伐他汀对 mDCs 具有一定的促凋亡作用。这些结果表明 FPP 增强了 CCR7 刺激的 mDCs 的线粒体呼吸作用和适应能力。
线粒体融合缓解了迁移性树突状细胞的内质网应激
下面研究 FPP 促进的线粒体融合是如何促进 CCR7 依赖性髓源性树突状细胞(mDC)的存活和迁移的。发现狼疮患者的浆细胞样树突状细胞(mig-DCs)中,内质网(ER)加工途径和氧化磷酸化途径显著富集。持续的 PERK(类蛋白激酶 R 的内质网激酶)和 p-eIF2α(磷酸化的真核起始因子 2ɑ)信号通路激活会导致内质网应激未解决以及促凋亡反应。在迁移中的树突状细胞中,线粒体融合是内质网应激减少的后续效应。从功能上讲,用线粒体分裂抑制剂 Mdivi-1 或线粒体融合促进剂 M1 处理髓源性树突状细胞(mDCs),可挽救在 Hmgcr 下调的 mDCs 中缺陷的 DC 存活,并且 M1 促进剂可挽救在 Hmgcr 缺陷的 mDCs 中减少的 mDC 迁移。总之,这些结果表明,FPP 促进的线粒体融合可缓解内质网应激,以维持 CCR7 介导的 DC 存活和迁移。
FPP 通过香叶酰基化促进 RhoA - MFN 相互作用
接下来探究了在 CCR7 刺激的髓源性树突状细胞中,将代谢物 FPP 与线粒体融合联系起来的分子机制。mTORC2 已被证明是细胞代谢的重要调节剂。尽管在 CCR7 刺激的髓源性树突状细胞中,FPP 上调了 mTORC2 的激活,但它并不影响 MFN2 的表达,也不介导 Hmgcr 依赖的髓源性树突状细胞迁移的促进。同时,在β-葡聚糖刺激的髓源性树突状细胞中,FPP 上调了 mTORC2 的激活,但不影响 MFN2 的表达。尽管 FPP 激活了 mTORC2,但 mTORC2 似乎并非在从 Hmgcr 依赖的线粒体融合到髓源性树突状细胞迁移的通路中发挥功能作用。FPP 能够直接或间接地调节蛋白质预基化,包括法尼基化和香叶基香叶基化,以调节细胞信号传导和行为。法尼基转移酶(FTase,由 Fnta 和 Fntb 编码)和香叶基香叶基转移酶(GGTase,由 Pggt1b 编码)分别催化法尼基和香叶基香叶基基团与以 CAAX 框(其中 C 为半胱氨酸,A 通常为脂肪族氨基酸,X 为任何氨基酸)结尾的蛋白质的羧基末端半胱氨酸的共价连接。GGTI 和 FTI 分别是 GGTase 和 FTase 的抑制性化学物质。我们发现,GGTI 显著抑制了 CCR7 触发的髓源性树突状细胞(mDC)迁移,但 FTI 没有这种效果。敲低 Pggt1b 会减少 mDC 的迁移,而敲低 Fnta/Fntb 则没有此类效果。GGTI 处理还消除了 CCR7 刺激的 mDC 中由 FPP 调节的线粒体极化。因此,FPP 通过蛋白质香叶基香叶基化而非法尼基化分支来促进线粒体健康和 mDC 迁移。蛋白质香叶酰基化已被证明能够调节诸如 Rho 和 Rab 蛋白等小 GTP 酶的功能。因此,我们进行了高效液相色谱 - 串联质谱分析。信号传导和细胞迁移。接下来,我们进行了综合筛选以寻找关键的预聚糖基化 GTP 酶靶点。在 25 个小型预聚糖基化 GTP 酶(分子量<25 千道尔顿)中,有 12 个候选物显示出 FPKM 值超过 100(反映高转录表达)(GSE184862)。对这些 12 个候选物进行有重点的 siRNA 筛选试验显示,RhoA 的敲低最显著地减少了 CCR7 触发的髓源性树突状细胞(mDC)迁移。因此,尽管 FPP 能够上调一系列小 GTP 酶在 mDC 中的预聚糖基化,但 RhoA 增强 mDC 迁移的能力最为显著。
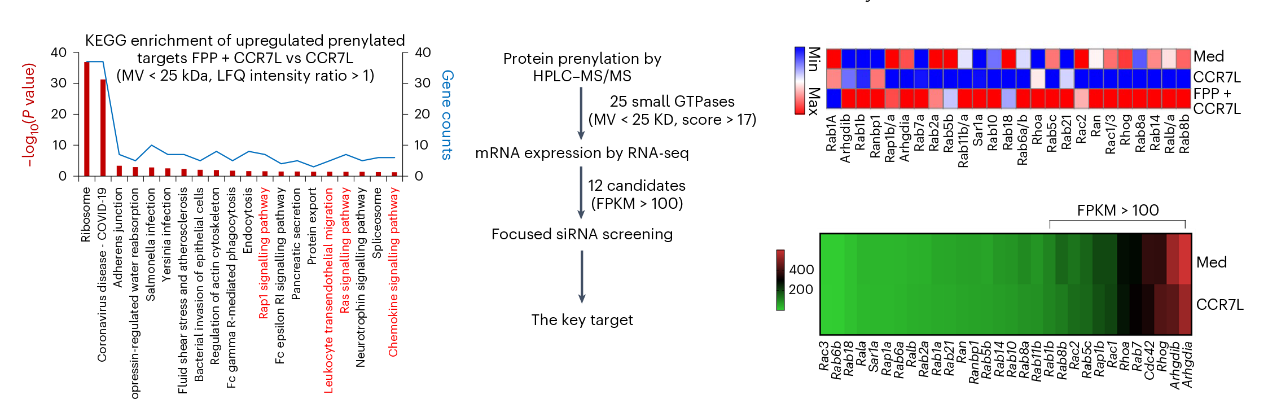
利用点击化学法,发现 Hmgcr 基因敲低降低了分子量约为 20 千道尔顿的蛋白质的香叶酰基化,FPP 通过介导 RhoA 香叶酰基化来促进 CCR7 介导的 mDCs 的存活和迁移。FPP 调节的 RhoA 线粒体定位促使我们进一步探究 RhoA 是否与 MFN1/2 结合。通过共免疫沉淀试验,我们表明 RhoA 与 MFN1 和 MFN2 结合,并且在 CCR7 刺激的髓源性树突状细胞中,FPP 刺激特异性地增强了 RhoA 与 MFN2 的相互作用。使用纯化的 GST-RhoA 和 His-MFN2 蛋白进行 GST 拉下实验进一步证实了 RhoA-MFN2 相互作用。这些数据表明,FPP 增强了 RhoA 与线粒体融合调节因子 MFN1/2 之间的相互作用。尽管 RhoA-G14V 的过表达增强了 RhoA/MFN2 相互作用,但 RhoA-G14V/C190R 的过表达阻断了这一过程。此外,RhoA-G14V 的过表达,而非 RhoA-G14V/C190R 的过表达显著缓解了涉及 PERK 和磷酸化 eIF2α 的促凋亡内质网应激通路的激活。总之,我们证明了 FPP 上调 RhoA 的香叶酰基化促进了线粒体融合,以促进内质网应激的缓解,从而增强了 CCR7 介导的髓源性树突状细胞的存活和迁移。
辛伐他汀可阻止树突状细胞迁移并预防狼疮自身免疫
接下来探究抑制胆固醇通路是否能保护自身免疫性疾病。给老鼠口服辛伐他汀(20 毫克/天),持续 3 周,然后让它们接受普里司他诱导的狼疮模型。发现辛伐他汀预处理 3 周可防止包括毛发在内的皮肤炎症损伤的发展。包括辛伐他汀、普伐他汀和洛伐他汀在内的三种不同的他汀类药物,均下调了 MRL/lpr 狼疮小鼠中IL-21+ T 细胞、迁移性树突状细胞、生发中心 B 细胞的比例。因此,Hmgcr-FPP 通路通过促进树突状细胞迁移及随后的生发中心反应而加剧狼疮自身免疫。与对照小鼠相比,我们从 SLE 小鼠分选的迁移性树突状细胞(mig-DCs)中检测到 MFN2 的表达增加,但内质网应激标志物 p-eIF2α 的表达减少,而辛伐他汀预处理可将其恢复至正常水平。自身免疫性疾病中迁移性树突状细胞中过度的线粒体融合和受限的内质网应激可以通过辛伐他汀恢复。
For mass spectrometry analysis of protein prenylation, mDCs, stimulated as indicated, were incubated for 12 h with 50 μM alkynyl-isoprenol chemical reporter mixed isomers. Cells were then lysed with RIPA buffer, obtained after ultrasonic disruption and subjected to the click chemistry reaction with CUAAC click labelling buffer for 1.5 h. Prenylated proteins were enriched by agarose streptavidin beads and subjected to liquid chromatography–mass spectrometry analysis on a Thermo Scientific Q Exactive mass spectrometer performed at AIMS Scientific Corporation (Shanghai, China), and raw mass spectrometry files were analysed by MaxQuant (v.2.1).



