蛋白序列分析
靶向代谢组实验专注于测定代谢物。三重四极杆质谱仪提供化合物确认所需的宽动态范围,高灵敏度以及出色选择性,是最佳选择。AIMS质谱提供专业的LC-MS和GC-MS服务,可为科研工作者提供下述代谢物质检测。
约 5–7 个工作日 空白 & 标准品 数据库检索 + 报告

靶向代谢组实验专注于测定代谢物。三重四极杆质谱仪提供化合物确认所需的宽动态范围,高灵敏度以及出色选择性,是最佳选择。AIMS质谱提供专业的LC-MS和GC-MS服务,可为科研工作者提供下述代谢物质检测。
约 5–7 个工作日 空白 & 标准品 数据库检索 + 报告
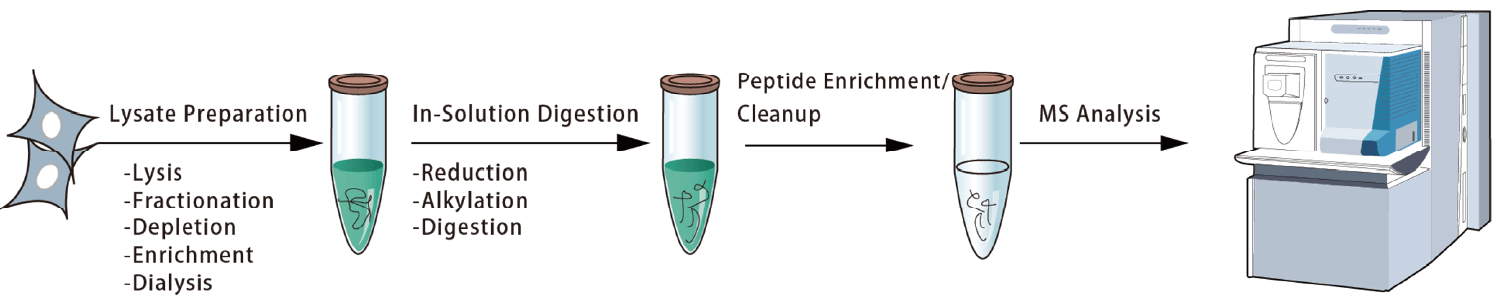
适用于细胞裂解液、免疫沉淀洗脱液以及缓冲液中的蛋白混合物。相较凝胶内酶解, 溶液法流程更简洁、肽段损失更少, 从而在更短时间内获得更深的蛋白组覆盖。

优化的还原/烷基化与酶切条件, 提升肽段鉴定数量。
优化的还原/烷基化与酶切条件, 提升肽段鉴定数量。
优化的还原/烷基化与酶切条件, 提升肽段鉴定数量。
优化的还原/烷基化与酶切条件, 提升肽段鉴定数量。
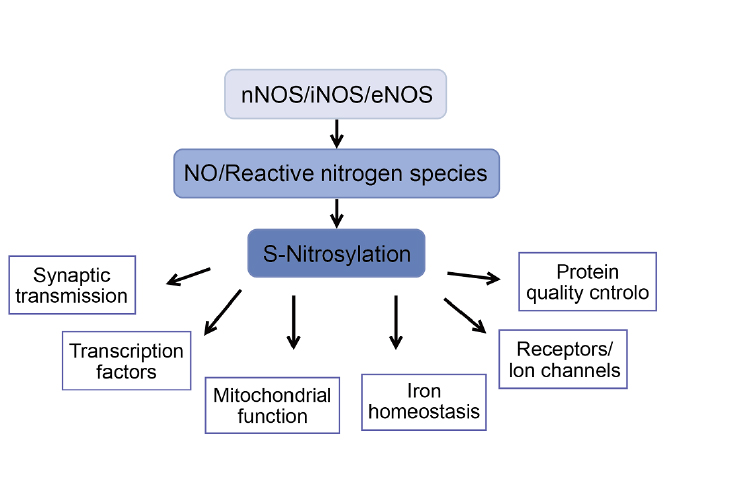
一氧化氮(NO)是诸如血管舒张, 血小板聚集抑制, 神经传递, 抗菌活动等一系列生理学过程的重要调控因子, NO的氧化还原状态及其二原子的化学性质, 使其能通过与各种蛋白质发生反应并调控各种细胞内和细胞外事件。很显然, NO修饰反应是通过将亚硝基集团转移至半胱氨酸的巯基上形成S-亚硝基硫醇(SNO), 这一过程通常被称之为S-亚硝基化作用。被大家公认的是蛋白发生S-亚硝基化的调控与磷酸化类似, 尽管蛋白中的半胱氨酸残基都可发生S-亚硝基化修饰, 但是只有很少的修饰是特异性的并且起到调控蛋白功能的作用。
目前最广泛阐明的NO信号通路机制包括对血红素蛋, 可溶性鸟苷酸环化酶以及细胞色素C氧化酶的结合和改变。蛋白S-亚硝基化是一种可逆的翻译后修饰, 被认为是蛋白质活动, 定位, 蛋白稳定性和蛋白之间互作的调节剂。
蛋白质的硫醇残基可以通过与NO集团和不同的硫醇底物发生特定反应形成S亚硝基硫醇, 其可以直接与NO作用也可通过在硫醇盐和S-亚硝基谷胱甘肽(GSNO)反应时形成。由于此过程的可逆性, 后一种化合物GSNO也同样促进S-亚硝基化反应, 因此会在氧化还原对GSH/GSNO和氧化还原对蛋白-SH/蛋白-SNO之间产生一种动态平衡。其实GSNOR通过与GSNO反应生成氧化谷胱甘肽二硫化物(GSSG)和氨(NH3), 因此而降低S-亚硝基醇的浓度, 随后谷氧还蛋白(GRX)通过将GSSG还原为GSH, 维护并稳定这个SNO的系统。 当然一氧化氮(NO)分子集团也可以直接转移到蛋白上发生修饰, S-亚硝基硫醇的动态平衡同样依靠于硫氧化还原蛋白/硫氧化还原蛋白还原酶(Trx/TrxR)系统, Trx-介导的SNO蛋白还原化为硫醇是通过对NAPDH的氧化为代价, 最终并产生NO去介导相关信号通路。
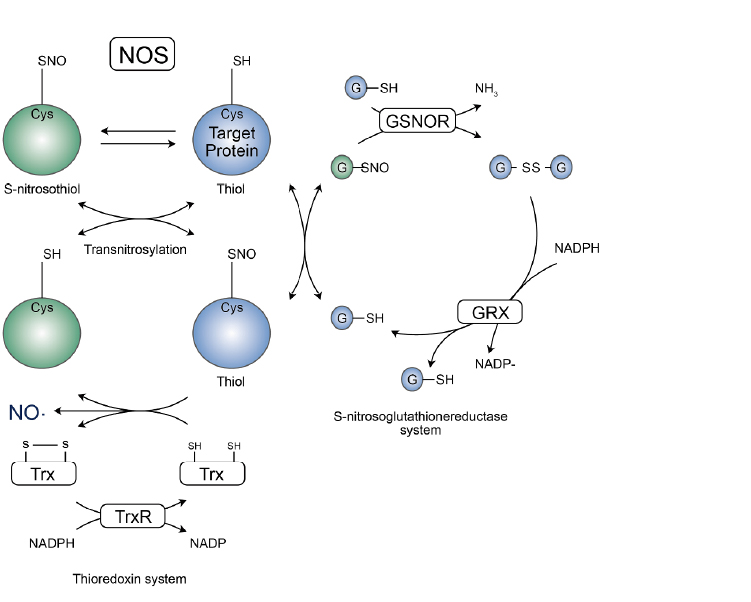
蛋白质的硫醇残基可以通过与NO集团和不同的硫醇底物发生特定反应形成S亚硝基硫醇, 其可以直接与NO作用也可通过在硫醇盐和S-亚硝基谷胱甘肽(GSNO)反应时形成。由于此过程的可逆性, 后一种化合物GSNO也同样促进S-亚硝基化反应, 因此会在氧化还原对GSH/GSNO和氧化还原对蛋白-SH/蛋白-SNO之间产生一种动态平衡。其实GSNOR通过与GSNO反应生成氧化谷胱甘肽二硫化物(GSSG)和氨(NH3), 因此而降低S-亚硝基醇的浓度, 随后谷氧还蛋白(GRX)通过将GSSG还原为GSH, 维护并稳定这个SNO的系统。 当然一氧化氮(NO)分子集团也可以直接转移到蛋白上发生修饰, S-亚硝基硫醇的动态平衡同样依靠于硫氧化还原蛋白/硫氧化还原蛋白还原酶(Trx/TrxR)系统, Trx-介导的SNO蛋白还原化为硫醇是通过对NAPDH的氧化为代价, 最终并产生NO去介导相关信号通路。
无标定量/谱计数 污染物过滤 支持自定义数据库
采用Thermo Fisher 高精度高分辨质谱仪, 可对蛋白序列进行精度检测, 上机时长120min更高的肽段覆盖率。
样本准备简单, 检测周期短, 约一周出报告。
若检测结果明显是质谱问题, 可免费重启上机一次。
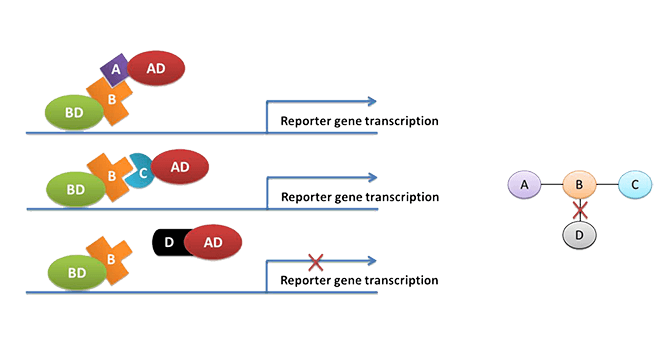
Advantages
用于高通量互相蛋白筛选 直接检测两个蛋白互相作用
2D
发现互作蛋白的数量有限
在实际细胞内部可能并不发生互相作用
不能发现蛋白非直接相互作用
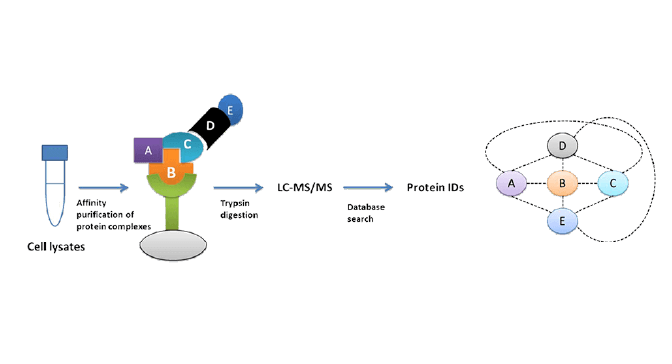
Advantages
用于高通量互相蛋白筛选 直接检测两个蛋白互相作用
2D
发现互作蛋白的数量有限
在实际细胞内部可能并不发生互相作用
不能发现蛋白非直接相互作用
采用 Thermo Fisher 高精度高分辨质谱仪, 直接对溶液样品进行酶解与鉴定, 流程简洁高效, 覆盖更深, 严格质控, 结果可直接用于科研发表。

解答实验需求与样品条件, 明确研究目标与预期结果。

根据需求制定实验方案, 确认检测范围、深度与交付内容。

客户提供溶液样品, 我们进行样品接收与质量初检。

执行溶液内酶解与质谱检测, 确保数据完整与可靠。

提供高质量检测报告, 数据符合科研发表标准。
查找有关试剂和一些常见使用规范常见问题解答, 如您找不到相关内容, 您可以了解我们在线客服.


